Prin Conducerea salată a excitației viteza de conducere suficient de rapidă a tractului nervos este asigurată pentru vertebrate. Potențialele de acțiune sar de la un inel neizolat la următorul pe axonii izolați. În bolile demielinizante, mielina izolatoare este descompusă, ceea ce perturbă conducerea excitației.
Care este conducta sărată a excitației?

Conducerea sărată a excitației este o formă de conductivitate nervoasă. În organismul vertebratelor, fibrele nervoase sunt izolate electric din împrejurimile lor de teaca de mielină și preiau astfel funcția unui cablu acoperit. Excitarea unei fibre nervoase apare la întreruperile acestui strat izolant, care sunt cunoscute și sub numele de inele de legătură sau noduri.
Multe fibre nervoase vertebrate au formă subțire. Axonii subțiri au o viteză de conducere mai mică decât tracturile nervoase puternice. Pentru ca viteza de conducere a nervilor să fie suficientă în ciuda rezistenței scăzute, conducția de excitație a vertebratelor este construită într-o manieră salată și folosește procese biochimice și bioelectrice pentru a transmite potențialele de acțiune.
Potențialul de acțiune sare dintr-un inel în celălalt în acest tip de conducere și lasă părțile învelite ale axonilor. Cu acest principiu se obține o viteză mai mare de conducere prin pompe de sodiu dependente de tensiune și procese biochimice bioelectrice.
Funcție și sarcină
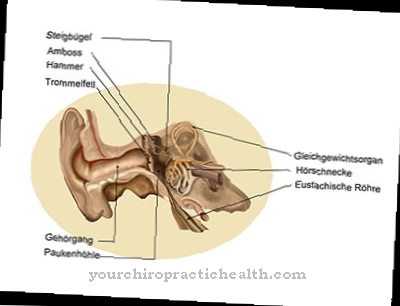
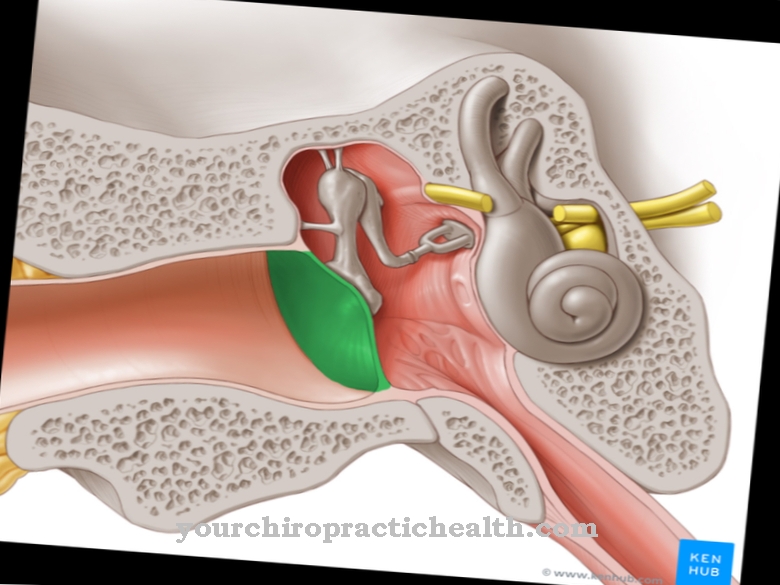
În sistemul nervos periferic, celulele lui Schwann formează mielina care învelește nervii. Oligodendrocitele își asumă această sarcină în sistemul nervos central. Axonii din ambele sisteme sunt acoperite cu mielină, care are un efect izolator electric. Izolarea axonilor este întreruptă la o distanță între 0,2 și 1,5 milimetri. Aceste întreruperi sunt, de asemenea, cunoscute sub numele de noduri sau legături Ranvier. În schimb, secțiunile acoperite cu mielină sunt numite internode și asigură o constantă de timp a membranei redusă, ceea ce asigură o viteză de conducere de 100 de metri pe secundă. Există, de asemenea, canale de sodiu + tensiune dependente în inelele de lacing fără teacă.
Atâta timp cât un axon nu este excitat, așa-numitul potențial de repaus prevalează în nodul său și de-a lungul internodului său. Între spațiul intracelular și spațiul extracelular al axonului există o diferență de potențial cu potențialul de repaus. Când un potențial de acțiune este generat pe primul con al liniei de excitație, care depolarizează membrana dincolo de potențialul său de prag, canalele Na + dependente de tensiune se deschid. Datorită proprietăților electrochimice, ionii Na + curg apoi din spațiul extracelular în spațiul intracelular.
Membrana plasmatică se depolarizează la nivelul inelului conului, iar condensatorul membranei este reîncărcat la 0,1 ms. În zona inelului de dantelă, există un exces intracelular de purtători de sarcină pozitivă în comparație cu mediul înconjurător din cauza ionilor de sodiu care au zburat. Este creat un câmp electric. Acest câmp generează o diferență de potențial de-a lungul axonului și are o influență asupra pieselor încărcate la distanța cea mai apropiată.
Particulele încărcate negativ pe următorul inel sunt atrase de excesul de încărcare pozitivă din primul inel. Particulele încărcate pozitiv între primul și al doilea inel de constricție se deplasează spre al doilea nod. Aceste schimbări de încărcare afectează pozitiv potențialul membranei celui de-al doilea inel con, deși ionii nu au atins-o. În acest fel, excitația sare de la inel la inel și păstrează proprietatea depolarizării suficiente a membranei inelelor ulterioare.
Boli și afecțiuni
Bolile demielinizante descompun teciile de mielină din jurul fibrelor nervoase. Aceste teci de mielină sunt o condiție necesară pentru conducerea salată a excitației. Fără teaca mielinei, pierderi mari de curent apar în internod. Prin urmare, sunt necesare excitații mai mari, astfel încât axonii să poată depolariza urmatoarele inele de formare cu un potențial de acțiune.
De regulă, potențialul de acțiune transmis după pierderi este prea mic pentru a fi recunoscut ca atare de următorul nod. Ca urmare, inelul de dantelă nu transmite emoția.
Fenomenul demielinizării este cunoscut și sub denumirea de demielinizare și aparține bolilor degenerative. Procesele legate de vârstă, precum și procesele toxice și inflamatorii pot marca marcarea axonilor și, astfel, punerea în pericol a transmiterii salate a potențialelor de acțiune.
Deficiențele de vitamine pot fi, de asemenea, legate de acest fenomen. Prea puțină vitamina B6 și vitamina B12 sunt asociate în special cu demarking. Un astfel de deficit de vitamine se găsește adesea în alcoolism, de exemplu. O demielinizare a sistemului nervos poate apărea și în contextul consumului de droguri.
Cea mai cunoscută cauză inflamatorie a decolorării nervilor este scleroza multiplă a bolii autoimune. Sistemul imunitar propriu distruge țesutul nervos din sistemul nervos central ca parte a bolii. Alte cauze ale demarcării pot fi diabetul, boala Lyme sau bolile genetice. Bolile genetice cu proprietăți demielinizante includ, de exemplu, boala Krabbe, boala Pelizaeus-Merzbacher și sindromul Déjérine-Sottas.
Simptomele care apar odată cu demielinizarea țesutului nervos depind de localizarea focarelor demielinizante. În sistemul nervos central, de exemplu, demielinizarea poate duce la o afectare a organelor senzoriale, în special la o afectare a ochilor. Paralizia este, de asemenea, concepută în cazul demielinizării în sistemul nervos central, deoarece tracturile nervoase motorii și centrele lor de control sunt situate acolo. În sistemul nervos periferic, demielinizarea nervilor este mai rar asociată cu paralizia. Pe de altă parte, demielinizarea axonilor periferici poate duce la amorțeală sau alte tulburări senzoriale.
Diagnosticul bolii demielinizante se face folosind tehnici imagistice precum imagistica prin rezonanță magnetică. Imaginile RMN prezintă, de obicei, focuri albe de demielinizare atunci când este administrat un agent de contrast.










.jpg)